朱正锦,陈 敏,田永贤,向贵生,张 颢,邱显钦
(1.云南大学 资源植物研究院,云南 昆明 650091;
2.云南省农业科学院 花卉研究所,云南 昆明 650205;
3.国家观赏园艺工程技术研究中心,云南 昆明 650205)
白粉病是切花月季种植中最严重的病害[1-3],白粉菌的掌状吸器吸取植株细胞中的营养物质维持自身的生长发育[4].大量研究发现MLO基因在植物抗白粉病中发挥重要作用,当MLO基因的碱基序列发生改变时会形成mlo等位基因,导致其蛋白功能丧失从而降低了MLO基因对白粉菌侵染的调控作用[4-6].将不同物种的MLO基因敲除后,发现大麦叶片出现斑点以及大麦谷粒产量下降[7-8];
拟南芥则表现出生长发育迟缓、衰败较早且未感染叶片有胼胝质沉积的现象[9];
辣椒表现为植株变小[10].本实验室基于大花香水月季与长尖叶蔷薇转录组数据,分别获得11和12个MLO基因,与其他物种比对,得到与响应白粉菌侵染相关的6个候选基因,其中符合抗白粉病MLO基因典型结构特征的有RgMLO6和RlMLO7基因[11].对6个候选基因进行qRT-PCR分析,结果显示RgMLO6和RlMLO7基因在受到白粉菌胁迫后相对表达量显著上调.
病毒诱导的基因沉默技术(VIGS)是一种能够快速简便对植物基因功能进行鉴定的遗传学方法.将目的基因片段导入病毒载体,然后侵染植物使该植物体内的基因被沉默,从而引起植株表型及生理指标发生改变,以此对目的基因的功能进行鉴定[12].目前,VIGS 技术已经被成功地应用于烟草(Nicotiana tabacumL.)[13-14]、拟南芥(Arabidopsis thaliana)[14]、棉 花(Gossypium spp)[15-16]、番 茄(Lycopersicon esculentumMill.)[17]、马铃薯(Solanum tuberosum)[18]等植物中.考虑到月季遗传转化的困难性,我们选择使用VIGS技术对RgMLO6和RlMLO7基因的功能进行初步鉴定.
本研究分析了大花香水月季的RgMLO6 基因和长尖叶蔷薇的RlMLO7基因在白粉菌侵染时的表达模式,进一步构建了该基因的VIGS沉默载体通过农杆菌介导侵染获得沉默植株并进行抗性鉴定.本研究为初步鉴定RgMLO6和RlMLO7基因对白粉病的调控功能,也为今后更好地分析MLO基因,培育蔷薇科植物抗白粉病新品种提供理论依据.
1.1 供试材料参照张颢等[19-20]的方法对来自大花香水月季与长尖叶蔷薇的幼叶进行白粉菌侵染,分别采摘未侵染(0 h)与侵染6、12、24 h的幼叶作为供试植物材料.接穗为云南省农业科学院花卉所科研成果示范基地的大花香水月季和长尖叶蔷薇萌发的腋芽,砧木为基地‘月月粉’(R.chinensis‘Pallida’)的枝条.大肠杆菌感受态细胞 DH5α 及根癌农杆菌热激感受态GV3101作为试验菌株,pTRV1和pTRV2两个供试载体.
1.2 试验方法
1.2.1RgMLO6和RlMLO7基 因 实 时 荧 光 定 量PCR 分别采摘白粉菌侵染0、6、12、24 h的大花香水月季与长尖叶蔷薇的幼叶,总RNA的提取与基因组DNA的去除使用植物总RNA提取试剂盒(北京天根生化科技有限公司).使用PrimeScriptTMRT reagent试剂盒(北京宝日医生物技术有限公司)将RNA反转录成cDNA.以在月季中稳定表达的UBC作为内参基因(表1)[2],采用qRT-PCR技术测定RgMLO6和RlMLO7基因的表达量,每个样品设置3个生物学重复[21].反应体系和反应程序参照向贵生[11]方法,采用2-ΔΔCt法[22]分析数据并用Origin 2021作图.
1.2.2RgMLO6和RlMLO7基因片段的克隆及表达载体的构建 总RNA提取和cDNA的合成同1.2.1节.分别选取RgMLO6基因CDS序列的3′端224 bp序列和RlMLO7基因CDS序列的3′端的259 bp序列,在设计引物(表1)时加上BamH Ⅰ和SmaⅠ酶切位点.PCR扩增产物经琼脂糖凝胶检测并回收目的条带,经测序验证后用于下一步试验.参照 TaKaRa公司试剂盒的操作步骤进行 PCR产物的切胶回收,将目的基因片段和病毒空载体pTRV2 同时用BamH Ⅰ和SmaⅠ进行双酶切[23].分别将酶切后的RgMLO6和RlMLO7片段与线性化载体pTRV2切胶回收.参照T4 DNA Ligase酶(TaKaRa公司)说明书得到连接产物,将连接产物转化至DH5α 感受态中,在含有50 μg/mL卡那霉素平板上挑取阳性单克隆,经菌液PCR检测后,进行测序鉴定.对测序鉴定正确的重组转化子菌液,按照 TIANprep Mini Plasmid Kit 说明书进行质粒提取[23],双酶切鉴定质粒是否携带目的基因.
1.2.3 农杆菌侵染及沉默效率检测 参照穆春等[24]的方法,通过电击转化法将质粒 pTRV2、重组质粒pTRV2-RgMLO6和pTRV2-RlMLO7转化至根癌农杆菌感态GV3101中,进行转化前的活化及重悬.分别取大花香水月季和长尖叶蔷薇长度在5 cm左右的腋芽置于清水中带回,参照晏慧君等[25]的方法进行抽真空侵染.将抽吸后的腋芽取出,放入清水中,通过劈接法进行嫁接,用封口膜将接口封住并套上袋子,每隔4 d观察接穗的情况.使用qRT-PCR分析接种20 d后的大花香水月季与长尖叶蔷薇的RgMLO6和RlMLO7基因的表达量,在RgMLO6和RlMLO7基因沉默片段外设计引物(表1),以UBS为内参基因(表1),扩增片段大小为109 bp.反应体系和反应程序参照Cao等[26]的方法进行,然后利用Origin 2021软件作图.

表1 本研究所用引物及其序列Tab.1 Primers used and their sequences used in the study
1.2.4 VIGS沉默RgMLO6和RlMLO7基因蔷薇科植株白粉病抗性鉴定 以VIGS沉默RgMLO6和RlMLO7基因的大花香水月季和长尖叶蔷薇作为处理组的实验材料,与对照组相比较进行白粉病的抗性鉴定.分别取白粉菌孢子悬浮液20 μL侵染处理组和对照组的6个嫩叶正面放入培养皿中密封,并置于20 ℃、光强3 000 lx、16 h/d的培养箱中培养7 d,然后统计嫩叶的白粉菌侵染情况,3次重复.基因沉默植株的抗病性依照张颢的方法进行判定[18],将相对抗病指数为1.00的判定为免疫(I)、在0.80~0.99之间的为高抗(HR)、在0.40~0.79之间的为中抗(MR)、在0.20~0.39之间的为中感(MS)、0.20以下的为高感(HS).
选用VIGS沉默RgMLO6和RlMLO7基因且生长状态相同的大花香水月季和长尖叶蔷薇(处理组)和未沉默的(对照组)的植株幼叶作为实验对象.分别取白粉菌孢子悬浮液20 μL侵染处理组和对照组的幼叶正面,并置于20 ℃、光强3 000 lx、16 h/d的培养箱中分别培养12、24 h和48 h,应用考马斯亮蓝染色法染色后在显微镜下观察白粉菌的生长状态.
2.1 RgMLO6和RlMLO7基因荧光定量qPCR以UBC为内参基因,对RgMLO6与RlMLO7基因在0、6、12 h和24 h时的相对表达量进行了分析.结果如图1所示,RgMLO6在6、24 h的表达水平分是未处理的24、11倍,差异明显,12 h的表达水平与未处理差异不明显;
RlMLO7基因在6、12 h的表达水平较是未处理的5、7倍,差异明显,24 h的表达水平与未处理差异不明显.这些结果表明大花香水月季的RgMLO6基因与长尖叶蔷薇的RlMLO7基因在白粉菌侵染中发挥重要作用.

图1 月季白粉菌诱导不同时期RgMLO6与RlMLO7基因的相对表达量Fig.1 The relative expression levels of RgMLO6 and RlMLO7 genes induced by powdery mildew rose at different stages
2.2 单克隆双酶切检测将RgMLO6和RlMLO7基因的CDS片段与线性pTRV2连接得到pTRV2-RgMLO6和pTRV2-RlMLO7重组表达载体,将重组表达载体利用BamH Ⅰ和SmaⅠ进行双酶切,最后菌液PCR验证正确性.结果如图2所示,PCR电泳得到2个272 bp和255 bp长度的小片段和一个长度接近线性载体pTRV2大小的片段,表明表达载体构建成功.
2.3 VIGS处理植株的荧光定量qPCR为了验证大花香水月季的RgMLO6与长尖叶蔷薇的RlMLO7 基因是否有效沉默,在VIGS载体转入20 d后进行荧光定量qPCR.如图3所示,结果表明沉默RgMLO6基因的大花香水月季中的RgMLO6基因表达量较对照组降低了87.5%;
沉默RlMLO7基因的长尖叶蔷薇中RlMLO7基因的表达量较对照组降低了88.9%.沉默植株中RgMLO6和RlMLO7基因的相对表达量显著下降了80%~90%,说明pTRV2-RgMLO6和 pTRV2-RlMLO7 的导入可显著地降低处理组植株内源目标基因的相对表达量,有效沉默目标基因.
2.4 VIGS技术处理后的2种植株的白粉病抗性鉴定为了进一步探究大花香水月季的RgMLO6与长尖叶蔷薇的RlMLO7 基因在抗白粉病中的作用,接种白粉菌至RgMLO6和RlMLO7基因沉默植株嫩叶7 d后对处理植株的白粉病抗性鉴定.如表2所示,经VIGS处理后大花香水月季的抗性水平由高感上升为中抗,长尖叶蔷薇的抗性水平由中抗上升为高抗,2种基因型植株的抗性水平均得到提高.
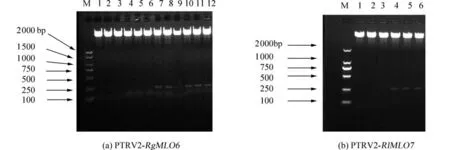
图2 双酶切检测载体重组表达载体Fig.2 Recombinant expression vector was detected by double enzyme digestion

图3 RgMLO6和RlMLO7基因在对照组和处理组植株中的相对表达量Fig.3 Relative expression of RgMLO6 and RlMLO7 gene in control and experimental plants

表2 对照组和基因沉默处理组植株白粉病抗性鉴定Tab.2 Identification of powdery mildew resistance in control group and gene silencing treatment group
经考马斯亮蓝染色后,显微镜观察对照组和RgMLO6和RlMLO7基因沉默处理组植株受白粉菌侵染后叶片中菌丝体的生长情况,结果如图4、5所示.大花香水月季和长尖叶蔷薇的叶表皮细胞在接种12 h后,对照组中均出现分生孢子和初级菌丝,而基因沉默处理组中仅观察到分生孢子.在接种24 h后大花香水月季对照组开始出现二级菌丝,长尖叶蔷薇在对照组中发现了分生孢子和较长的初级菌丝,RlMLO7基因沉默处理组中发现分生孢子和初级菌丝.在接种48 h后对照组发现了串生分生孢子,而在基因沉默处理组中仅发现了分生孢子和初级菌丝.
MLO基因具有负向调控植物抗病性和叶肉细胞死亡双重功能[27-29].邱显钦等[2,6]从野蔷薇中克隆得到4个mlo基因并进行结构、亚细胞定位及时空表达模式分析,克隆1个MLO基因RmMLO并鉴别出该基因在月季相应白粉菌侵染中有一定作用.Kaufmann等[30]将月季中MLO家族成员进行分析,筛选出4个与月季白粉病互作的MLO候选基因(RhMLO1~RhMLO4).随后Qiu等[31]对RhMLO1-RhMLO4基因进行qPCR分析,结果发现在受到白粉病菌侵染不同阶段表达量均明显上调的有RhMLO1和RhMLO2基因.Qiu等[32]通过PCR和Southern blot检测转基因株系的整合和拷贝数,使用实时荧光定量PCR和抗性分析发现野蔷薇的抗性与沉默RhMLO1的积累有明显的相关性.本研究通过qRT-PCR分析RgMLO6 和RlMLO7基因在受到白粉菌侵染0、6、12、24 h的表达量,结果表明与对照相比不同时期的表达量显著上调,这与在葡萄、西瓜中的研究结果相一致[33-34].

图4 白粉菌在大花香水月季幼叶上菌丝体生长情况Fig.4 Mycelial growth of powdery mildew fungi on young leaves of Rosa odorata var.gigantea

图5 白粉菌在长尖叶蔷薇幼叶上菌丝体生长情况Fig.5 Mycelial growth of powdery mildew fungi on young leaves of Rose.longicuspis var.longicuspis
应用VIGS技术沉默大花香水月季和长尖叶蔷薇RgMLO6和RlMLO7基因,结果发现沉默表达载体导入后有效抑制了RgMLO6和RlMLO基因的表达,表明VIGS技术沉默效果明显.对RgMLO6和RlMLO7基因沉默植株(处理组)和对照组植株进行白粉病抗性水平鉴定,结果表明较对照组处理组植株的抗性水平都得到提高.显微镜观察对照组和RgMLO6和RlMLO7基因沉默(处理组)植株叶片在受到白粉菌侵染后菌丝体的生长情况,整体表现出当植株受到白粉菌侵染后基因沉默植株中菌丝体的生长较对照组更慢.这些结果均表明当植株内源MLO基因的表达下调时,可以有效提高蔷薇科植物对白粉病的抗性.
RgMLO6和RlMLO7基因参与了寄主与白粉菌的互作过程,对蔷薇科植物白粉病起负调控作用.
猜你喜欢白粉蔷薇白粉病蔬菜白粉虱 秋防正当时今日农业(2021年19期)2022-01-12一到春季就流行 蔬菜白粉病该咋防今日农业(2021年9期)2021-11-26蔷薇满架,好眠一夏基层中医药(2020年6期)2020-09-11花开恣意是蔷薇人大建设(2018年7期)2018-09-19保护地蔬菜白粉虱的发生与防治新农业(2017年2期)2017-11-06拉萨设施月季白粉病的发生与防治西藏科技(2016年8期)2016-09-26用于黄瓜白粉病抗性鉴定的InDel标记中国蔬菜(2015年9期)2015-12-21白僵菌防治温室大棚白粉虱药效试验生物灾害科学(2015年3期)2015-03-11黄瓜白粉病的发生与防治现代农业(2015年5期)2015-02-28蔷薇白发抄火花(2015年7期)2015-02-27